
Introduction
Conservation biology benefits from the inclusion of historical interpretations to ensure that its assessments are comprehensive and its proscriptions remain robust (Meine 1999; Szabo and Hedl 2011). One challenge faced by practitioners concerns back-casting to estimate the onset of deleterious change (Novaglio et al. 2020), as a way to counter to the ‘shifting baseline syndrome’ (see Pauly 1995 and Jackson, Alexander & Sala 2011), wherein successive generations become accustomed to regard an ever-diminishing biodiversity as being ‘natural.’ The work of social scientists (Ellis et al. 2016), and particularly environmental historians (Dearing et al. 2015), is critical in this regard through searching out and interpreting social-ecological information buried within archival documents. For example, historical ecologists involved in understanding anthropogenic change in marine ecosystems (McClenachan, Ferretti & Baum 2012; Kittinger et al. 2015) have championed the view that insights pertinent to conservation biology can be obtained from non-traditional sources unrelated to commercial fisheries, such as first-person fishers’ observations, travellers’ diaries, explorers’ reports, naturalists’ journals and ethnographic records (e.g. Saenz-Arroyo et al. 2006; Al-Abdulrazzak et al. 2012). The present study uses this approach of mining historical narratives to provide anecdotal evidence for the antecedent entanglement of marine fauna in African waters.1
Garbage now completely encircles the planet, with even the remotest corners far removed from human inhabitation containing the copious flotsam of our human existence. This has led some to propose that ubiquitous waste be considered one of the defining features of the Anthropocene (Hecht 2018). Indeed, there are few more alarming prognostications than that, within a generation, the mass of discarded plastic littering the oceans will exceed the biomass of all marine life (Kaplan 2016). One very real environmental threat of the Anthropocene is the entanglement of marine animals in plastic fishing gear, something which is known to impact more than two hundred species worldwide (Laist 1997). In terms of the thesis developed in this paper, it is important to note that despite ensuing debilitations resulting from infected wounds and impeded mobility caused by rope abrasion and hydrostatic drag (Derraik 2002), nonlethally entangled animals are known to pull trains of entangled debris over distances of thousands of kilometres and for durations of months and sometimes years (e.g. Johnson 2005; Neilson et al. 2009; Anon. 2019).
A question for historical ecologists interested in probing the thorny issue of the shifting baseline syndrome is when entanglement became a threat to the wellbeing of marine fauna. Due to the assumption that natural materials will ‘lose their resilience in usage and if lost or discarded at sea [will] tend to disintegrate quickly’ (Gregory 2009), entanglement is widely considered by conservation biologists to be a recent phenomenon corresponding to the advent and widespread use of plastic since the middle of the twentieth century, with little or no occurrence before that time (NOAA 2014; Vegter et al. 2014). In point of fact, hemp, flax, and cotton ropes and nets – often impregnated with tar or soaked in tanning solutions – were of sufficient durability (Aiken and Purser 1936; McCaskill 2009) to justify their continued and widespread maritime use (Bekker-Nielsen and Casola 2001). Because of this, they posed an entanglement threat. Furthermore, because natural fibre nets were neither as strong nor as elastic as their modern equivalents made of synthetic material, a higher proportion of animals would have evaded capture, making off with portions of the net and its accompanying floats in tow. And in so doing, an opportunity would have been created for such entangled animals to be observed and misconstrued as what cryptozoologists customarily refer to as the ‘many-humped’ or ‘string-of-buoys’ category of sea monster (Heuvelmans 1968). In particular, net-floats during the nineteenth, and part of the twentieth, centuries were made from blown glass balls, pieces of cork, and wooden casks. Observing such a train of such buoys (Figure 1), bobbing up and down on the surface of the water while being pulled behind a swimming animal and possibly festooned with strands of seaweed, it is easy to imagine how eyewitnesses might have been misled into believing that the entangled fishing gear or hunting equipment was the long tail of a sea serpent. Such a belief would have certainly been within the prevailing zeitgeist.

Strings of fishing-net floats which, if nonlethally entangled around an actively swimming animal and bobbing up and down on the water surface might be misinterpreted as the long tail of a presumed sea serpent, especially if the UMO was also pulling a ‘mane’ of intertwined fishing net and accumulations of seaweed or other natural or anthropogenic debris. Left image: cork pieces used to buoy a gillnet (photo taken at the Cape Ann Museum, MA, USA; from France 2019a); Right image: casks used to suspend a purse-seine (Goode et al. 1884; reproduced in France 2019a).
During the nineteenth century, many of the world’s leading natural scientists (see Regal 2012) commented upon sea serpents/monsters in dozens of articles published in the leading scientific journals (Westrum 1979). Eyewitness accounts of sea serpents therefore provide insights about the social and ethno-zoological history of human-nature relationships (e.g. Brown 1990; Lyons 2009; France 2019a). Today, in consequence of greater knowledge about marine fauna and optical physics, scientists have posited known species and advanced discernable phenomena to explain many early sightings of unidentified marine objects, or UMOs (e.g. Lehn and Schroeder 2004; Paxton, Knatterud and Hedley 2005; Galbreath 2015). Recently, there has been recognition that some sightings of imaginary sea serpents actually provide evidence for real environmental threats (France 2017). Putative sea serpents, reinterpreted in this fashion, represent monstrous acts of the Anthropocene rather than monstrous beings from natural history, and in consequence, are better examined through a lens of historical conservation biology rather than modern cryptozoological fantasy (France 2020).
Compilation of historical accounts from around the world for what were considered to be sea monsters reveals five regional clusters of abundant sightings (Heuvelmans 1968). Deductions have been made that a proportion of the sightings for three of the regions – the Western Atlantic, the Western Pacific, and the British Isles – can be parsimoniously explained (as per Das 2009) as unrecognised animals that were nonlethally entangled (France 2020 and references therein). The present paper extends the investigation to consider another of those so-called monster-laden hotspots: Africa (particularly, but not exclusively, the southern portion of the continent). Two sightings of UMOs from this region have previously been proposed, based on information contained in the eyewitness descriptions and illustrations, to have been animals pulling anthropogenic debris: the 1848 sighting between the Cape of Good Hope and St Helena by the crew of HMS Daedalus (de Camp and Crook de Camp 1985), and the 1857 sighting from the Cape Town shore by a number of citizens (Heuvelmans 1968). The purpose of the present investigation was to expand the examination to include the entire corpus of UMO sightings surrounding the African continent made over a period of two centuries, in order to determine if those two sightings were atypical or whether there is evidence for the widespread, pre-plastic entanglement of marine fauna in circum-African waters. If the latter, this would contribute to the growing evidence that entanglement has a much longer environmental history in the Anthropocene than is commonly believed, and supports the illation that historic sightings of ‘sea monsters’ can provide useful information on important issues for modern conservation biology (Parsons 2004).
The focus on Africa is significant in this regard since, with an environmental history as rich and as troubling as anywhere else (e.g. Beinart 2000; Carruthers 2004), it holds a fascinating position in discussions about the Anthropocene. Largely this arises in response to the contention of geographical determinism wherein the continent’s suffering from rampant disease has always placed it at a disadvantage (Diamond 2007). In point of fact, Africa’s development, despite its location and centuries of colonial interference, is a remarkable feat (Akyeampong et al. 2014). In consequence, impressions of backwardness are misplaced (to be kind), such that the continent should be viewed as an active and culpable participant in Anthropocenic change (Hecht 2018).
Methods
The Study Area
To Victorians, sub-Saharan Africa was ‘the Dark Continent’ (e.g. Stanley’s Through the Dark Continent and In Darkest Africa, and Conrad’s The Heart of Darkness; Anderson 2016). Contrary to popular belief, this was not in specific reference to the skin colour of its human inhabitants but nevertheless did reflect the implicitly racist perception about those peoples’ imagined savagery and barbaric behaviour, which was used to justify the imperial and missionary underpinnings of the ‘Scramble for Africa’ period of colonialisation. But ‘dark’ also referred to the fact that for nineteenth-century Europeans and North Americans, the interior of the African continent remained largely unknown; a blank on the map that was believed to be filled with all manner of surprising dangers and sinister mysteries; a place where, since classical antiquity, fearsome animals and monstrous races were thought to roam undiscovered. And it is in the same spirit of equating ‘darkness’ with poor understanding that led Pimm (2007) to use the analogy to describe the state of knowledge about modern conservation biology on the continent.
Today, biologists recognise Africa to be a biodiversity hotspot (France and Rigg 1998; Melles et al. 2019) that contains the largest number of megafauna species due to being the continent least impacted by the Pleistocene extinctions. Because medieval Europeans considered Africa to be one of the most likely candidate locations for the Garden of Eden (Relaño 2004), it achieved a near mythical status (Brantlinger 1985; Jarosz 1992) as a place harbouring many species, some of which originated from before the Fall. Because of this, Europeans had long been fascinated by exotic African animals (e.g. Lloyd 1971; Belozerskaya 2009; Grigson 2018), and the thought that there might be other, as yet unknown, species just waiting to be discovered by intrepid Victorian adventurers was a most intriguing possibility. This held a compelling allure (Miller 2012) since, in the common wisdom of the time, strange chimera and monstrous creatures were thought to inhabit the edges of the known world, including, most notably, Africa.2
Present-day marine fauna in Africa are known to be susceptible to entanglement and becoming incidental by-catch (e.g. Cliff et al. 2002; Razafindrakoto et al. 2008; Meyer et al. 2011), something which, as found elsewhere, is thought to be due to the modern use of plastic. However, given that fishing equipment has been deployed within the continent’s coastal waters for millennia (Brewer and Friedman 1989; van Niekerk 2011; Fagan 2017), it is my contention that such threats to marine life are unlikely to be a new phenomenon. Indeed, if Alverson et al. (1994) and a senior NOAA scientist (interviewed in Deedy 2017) are correct, and that nonlethal entanglement has existed ever since humans first threw spears and floats into the waves and set nets and traps in the water, then it should be possible to detect its presence hidden within the words and in the illustrations of historical sightings of misconstrued sea serpents. Certainly, the African coast during the nineteenth century would, just as everywhere else in the world at the time, have provided an obstacle course of anthropogenic material – in this case, originating from artisanal fisheries by African peoples and possibly European colonisers – that would have posed a notable entanglement hazard to marine life.
The Precedent for Comparison
The precedent that sightings of fantastical monsters in African waters were really mundane animals entangled in fishery equipment that went unrecognised at the time, derives from the case of New England’s nineteenth-century ‘Gloucester Sea Serpent,’ the most sighted and studied such creature in history. The evidence for such a contention is compelling (Fama 2012; France 2019a), as for example the ‘duck test’ of abductive reasoning wherein dozens of eyewitnesses described the UMO’s body as resembling a string of fishing-net buoys floating upon the surface of the water. For, as Loxton and Prothero (2015: 233) state in their polemic against cryptozoology, ‘a humungous serpentine animal might resemble a string of buoys, but a group of smaller individual objects (say, an actual string of buoys) also might resemble a string of buoys’ (my italics). Another dozen morphological and behavioural attributes characteristic of the Gloucester UMO have also been used as evidence for conclusions made of the prevalence of early, pre-plastic entanglement of megafauna in other regions (France 2020).
The Anecdotal Provenance
The historical source that was mined for anecdotes is Heuvelmans’ (1968) seminal book, recognised by scholars for its comprehensive listing of sea serpent sightings compiled from hundreds of disparate sources, including newspaper reports, magazine articles, legal documents and scientific papers, which was built upon the earlier, and similarly assembled, compendium by Oudemans (1892). A sub-set of African sightings, representing all nine of Heuvelmans’ (1968) categories of putative sea monsters observed between 1829 and 1949 were found to be of sufficient detail to enable their careful parsing through directed content analysis (Hsieh and Shannon 2005) with grounded coding (Charmaz 2007) based on six physical and seven behavioural attributes or traits that had been specifically identified as characteristics of the entangled Gloucester UMO. From the complete corpus of 50 examined sightings, 29 accounts (58%) suggested the observed UMOs to have been nonlethally entangled animals. About half of the encounters occurred while ships were ‘rounding the Cape’ in transit from Britain to colonial India. Table 1 documents locations for these anecdotal sightings: further details about the specific sightings can be obtained from their extended descriptions in the diachronic listings in both Heuvelmans (1968) and Oudemans (1892).
Sighting locations of African unidentified marine objects (UMOs) observed in pre-plastic times (1829–1949). See Oudemans (1892) and Heuvelmans (1968) for further details.
| (1) | 1829; near Cape of Good Hope (henceforth the ‘Cape’), South Africa; widespread referral to by settlers. |
| (2) | 1845; Cape Town, South Africa; numerous witnesses. |
| (3) | 1848, South Atlantic between the Cape and St Helena; famous sighting from HMS Daedalus (Galbreath 2015). |
| (4) | 1852; south of Mozambique. |
| (5) | 1852; west-central coast of continent. |
| (6) | 1854; South Atlantic off St Helena. |
| (7) | 1854; rounding the Cape. |
| (8) | 1856; near the Cape; ship Princess. |
| (9) | 1857; off Cape Town (France 2018). |
| (10) | 1857; northeast of St Helena; the crew. |
| (11) | 1858; between the Cape and St Helena. |
| (12) | 1863; between the Canary and Cape Verde islands. |
| (13) | 1871; near Durban, South Africa. |
| (14) | 1878; Gulf of Aden, Somalia; steamer Poonah. |
| (15) | 1881; near Cape Town; number of people on shore. |
| (16) | 1883; off Libreville, Gabon. |
| (17) | 1884; off Durban. |
| (18) | 1886; north of Durban. |
| (19) | 1886; Red Sea, Egypt. |
| (20) | 1889; Madagascar. |
| (21) | 1889; off Algiers, Algeria. |
| (22) | 1893; Namibia coast. |
| (23) | 1899; Algeria coast. |
| (24) | 1903; near Cape Town. |
| (25) | 1910; Mozambique Channel. |
| (26) | 1912; near Port Elizabeth, South Africa. |
| (27) | 1913; Gulf of Aden. |
| (28) | 1934; Somalia coast. |
| (29) | 1948–49; off Mombasa, Kenya. |
Results
In toto, a notable anatomical feature of the examined African UMOs (Table 2) is their considerable length, ascribed by eyewitnesses to various sizes, including being less than ten metres (one anecdote), between ten and 20 metres (seven anecdotes), between 23 and 40 metres (six anecdotes), to more than 40 metres (five anecdotes), and in two cases, to even more than 100 metres. Bodies are frequently referred to as elongated and narrow, described as being serpentine or snake/eel-like in form (16 anecdotes), with the occasional mention (two anecdotes) made of an absence of fins. Tellingly, and most notably, much of the lengths of the UMOs are composed of overt series of irregular or uneven body parts such as ‘knobs’ in the form of a dorsal ‘ridge’ or ‘crest’ (eight anecdotes), that are also described as ‘arches’ or ‘coils’ (three anecdotes) or likened to resembling a rope of fishing net floats (three anecdotes), sometimes accompanied by numerous lateral projections thought to be ‘fins’ (two anecdotes). Bodily features are occasionally identified as resembling casks or scales (eight anecdotes), with the noted presence of variously described accoutrements (seven anecdotes), including, significantly, the conspicuous occurrence of long strands of material, often interpreted as ‘hair’ or a ‘mane’, that sometimes looks like filamentous seaweed (eight anecdotes).
Descriptions of anatomical attributes of UMOs, imagined to be sea monsters, observed between 1829 and 1949 in African waters, and displaying traits synonymous to the Gloucester Sea Serpent and indicative of entangled fauna (France 2019a). Anecdotes 1 to 29 are indicated in brackets. See Oudemans (1892) and Heuvelmans (1968) for further details.
| Notable length | ||
|---|---|---|
|
||
| Narrow, tapering, sinuous, snake/eel-like shape, sometimes with absence of a head, and often with no caudal fin or lateral appendages | ||
|
||
| Body composed of a series of irregular, jointed component parts (multiple humps, coils, fins or a ridge) | ||
|
||
| Body components likened to kegs or barrels, and sometimes to shiny scales or saucers | ||
|
||
| Overall body likened to a string of floats, kegs or buoys | ||
|
||
| Presence of a horn, spike, spine, mane of hair resembling seaweed, or other debris protuberance (sometimes identified as a head) | ||
|
||

Two African UMOs that were described as sporting multiple dorsal ‘fins.’ Left image: the 1856 Princess sighting from South Africa (Anecdote 8). Right image: the 1878 Poonah sighting from Somalia (Anecdote 14). Obviously, megafauna do not exist which have propulsive fins on their backs held above the surface of the water and which angle forward against the direction of swimming movement. In consequence, rather than these being ‘cryptids’ completely new to science, a more parsimonious explanation is that they are unrecognised animals pulling series of net-floats or other segmented debris. Images from the Illustrated London News.
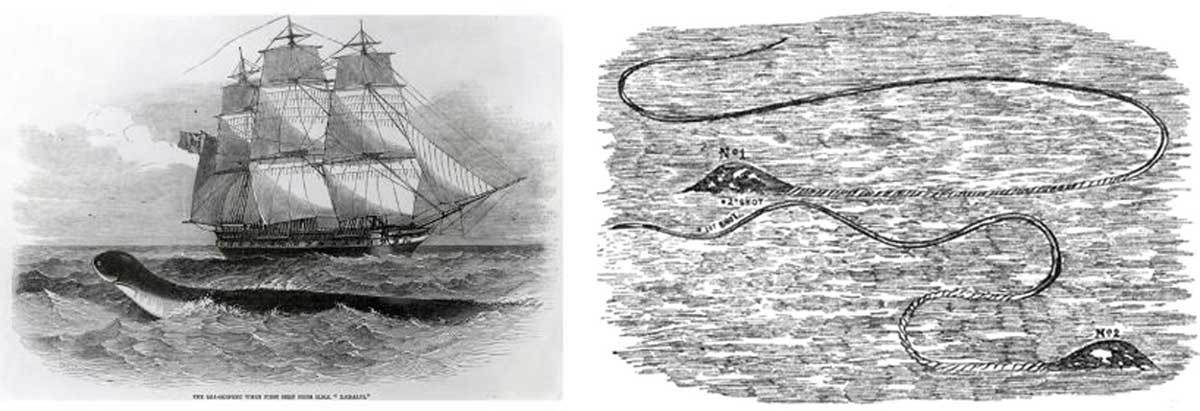
Purported exotic sea serpents of the mysterious ‘Dark Continent’ or simply mundane marine animals from Africa trailing entangled fishing gear or maritime debris? Left panel: the 1848 Daedalus sighting between St Helena and South Africa (Anecdote 3), hypothesizsed to have been a dugout canoe towed by a submerged whale shark (de Camp and Crook de Camp 1985). Right panel: the 1857 sighting off Cape Town (Anecdote 9), implied (Oudemans 1892) and explicitly suggested (Heuvelmans 1968; France 2018) to have been a surface-swimming mammal pulling a fishing rope (note the braided nature of the ‘tail’). Images from the Illustrated London News.
A notable behavioural feature of the African UMOs (Table 3) is their rapid speed of movement (seven anecdotes), which is accompanied by considerable water disturbance (six anecdotes). Observations reveal that in one case the elongated UMO displayed itself on the surface of the water with a high degree of flexibility, and most notably, for six cases, their body components were observed to move independently through vertical undulations. When not swimming slowly or floating motionless upon the surface (eight anecdotes), the extended bodies of UMOs were sometimes seen to suddenly be pulled beneath the waves (four anecdotes), or, in one case, to thrash about as if in distress. Portions of UMOs observed above the surface seemed oblivious to their surroundings (three anecdotes), including one case of passing right underneath a ship and another of remarkably even failing to respond after being struck by rifle shot.
Descriptions of behavioural attributes of UMOs, imagined to be sea monsters, observed between 1829 and 1949 in African waters, and displaying traits synonymous to the Gloucester Sea Serpent and indicative of entangled fauna (France 2019a). Anecdotes 1 to 29 are indicated in brackets. See Oudemans (1892) and Heuvelmans (1968) for further details.
| Rapid speed of movement |
|---|
|
| Obvious trailing wake or water disturbance |
|
| Notable flexibility of body |
|
| Vertical undulating movement of body segments |
|
| Floating, sometimes motionless, gently swaying in waves, or moving very slowly |
|
| Extended body pulled down into water, thrown up into the air, or thrashing about on the surface |
|
| Oblivious of surroundings or impervious to disturbance |
|
Discussion
From Imaginary Monsters to Real Monstrous Acts: Entanglement Interpretations
This paper is not the first time that ‘sea serpents/monsters’ swimming about Africa have been interpreted as being mundane animals entangled in fishing gear, hunting equipment, or other maritime debris. Excluding the Gloucester Sea Serpent (Brown 1990; France 2019a) and Egede’s ‘most horrible monster’ (Paxton et al. 2005; France in press), no other sighting of a singular UMO has elicited as much global discussion and debate as the 1848 Daedalus creature (Anecdote 3, Figure 3). Leading Victorian naturalists, including Sir Richard Owen (Regal 2012), commented upon the nature of the UMO in a flurry of journal articles (Lyons 2009). Parsimonious theories began almost immediately with the sighting of another putative sea serpent in the same area just a few weeks later that was ‘covered with a long shaggy-looking kind of mane’ (Heuvelmans 1968: 201). Remarkably, in this particular case, it was not until a dory had been launched and the ‘monster [which had been] all the time ducking its head, and showing its great length’ was captured and hoisted on board the ship ‘before it was discovered to be a piece of gigantic seaweed…the root end of which appeared when in the water like the head of the animal, and the motion given by the sea caused it to seem alive’ (this shows just how easy it is to be fooled, even for experienced mariners). The fascination with the Daedalus Sea Serpent has persisted until today, with Galbreath (2015) most recently positing the UMO to have been a surface-skimming roqual whale. de Camp and Crook de Camp (1985: 287–288), however, offer an alternative theory:
If one looks at the Illustrated London News picture squint-eyed, one sees that the ‘serpent’ looks like a dugout canoe, of the kind used in primitive lands for fishing. The dark colour of the ‘serpent’s’ back would be the shadowed interior of the canoe, and the ‘head’ the expanded bow platform on which a fisherman places his foot in shooting or spearing a fish.
Our surmise, then, is that some fishermen, fishing from their dugout off the coast of West Africa or eastern South America, rashly harpooned a whale shark [Rhincodon typus] … The fish, naturally, took off. The harpoon line was belayed to the canoe. Failing to untie or cut the line, the fishermen dove overboard and swam for shore, leaving the shark to tow the boat about the South Atlantic for weeks or months until the line broke or the harpoon head tore out.
There is a precedent for such a conclusion, given that whales and large fish such as sharks and tuna have been hunted with thrown harpoons for millennia (Fagan 2017). In a process referred to as ‘kegging’, a series of floats (sometimes made of animal bladders), and occasionally even a small boat, would be attached to the harpoon line in order to create enough drag to slow down the struck creature. In pre-ballistic times, fully one quarter of all struck animals avoided capture (Mowat 1997; Gardner 2007). In one famous case, for example, the polar explorer Nansen lost three ropes and their accompanying strings of floats to an escaping whale (described in Mowat 1997). Even today, whale sharks are found with harpoons embedded in their flesh, a legacy of their near brush with death (Riley, Harman and Rees 2009). And it is not unknown for struck megafauna to even make off with ropes attached to the small fishing craft from which the hunters had hurled their harpoons. For example, Herre (1942) describes hunting by native islanders in the Seychelles: ‘Sometimes when a whale shark is harpooned it dives with a very great rapidity until the rope is all out, and then keeps on downward until the boat [a dugout canoe] is carried under before the crew has time to escape.’
Secondly, with respect to the illustration (Figure 3) of the 1857 Cape Town sighting (Anecdote 9), Oudemans (1892: 237) comments that ‘nobody can help laughing when he sees this figure, representing something very much like a black buoy, with white streaks and spots, and glittering in the sun, having a long rope attached to it!’ Notably, he also remarks upon the ‘astonishing lateral flexibility’ of the UMO that closely matches that described previously for the Gloucester Sea Serpent, now recognized to have almost certainly been an entangled animal, possibly a whale (Fama 2012), but more likely a large fish like a giant bluefin tuna (France 2019a). Heuvelmans (1968: 242) is in agreement:
Dr. Biccard’s picture shows something more like an immensely elongated tadpole or a gigantic spermatozoon than a serpent. One wonders why he called the only substantial part of it a ‘head’, when it looks more like a ‘body’, and the long twisted tail, that looks nothing so much as a floating length of coir rope, the ‘body’. This so-called body is so unlike any part of an animal that one cannot help thinking that it may have been a net or rope towed by a shark or porpoise which had got caught in it and whose wounded body appeared to be what the doctor called the head.
As remarked upon previously (France 2018), from an historical perspective, it is interesting to note this statement of Heuvelmans’ was made in the 1960s at a time when the threat posed by entanglement had yet to be appreciated by marine conservation biologists.
The present archival examination of the complete corpus of sea serpent sightings from circum-African waters, made when the landmass was regarded as the mysterious ‘Dark Continent’ and thought to harbour all manner of undiscovered beasts, suggests that putatively entangled animals were a widely observed phenomena. The prodigious bulk of the 30m-long blue whale (Balaenoptera musculus), the largest animal known to have existed, and the miniscule girth of the 55m-long bootlace worm (Lineus longissimus), the longest animal alive today, removes their candidacy as misconstrued African sea serpents. Moreover, the considerable lengths reported for most of the African UMOs are, with the exception of whale sharks and a handful of cetacean species, counter to the body sizes of marine fauna (Freedman and Noakes 2002; McClain et al. 2015) known to inhabit the seas surrounding the continent. Due to their body proportions and breathing behaviour, whales, when associated with UMO sightings, are generally recognised (France 2016). In contrast, for these African UMOs, it is obvious that they must be preternatural, as there are no animals of such comparable length that have narrow dimensions like those of snakes or eels. As shown in Table 4, these observed attributes of UMOs from around Africa are in agreement with those of the Gloucester UMO (France 2019a) and others from the British Isles, North America and the Western Pacific (France 2020) proposed to have been entangled animals. This concordance across different regions gives credence to the suggestion of a consistent explanation.
Observed physical and behavioural attributes of UMOs posited to have been entangled animals. Attributes are those used to describe the UMO observed in and around Gloucester in 1817 and clearly indicative of an animal entangled in marine debris (France 2019a). Numbers shown indicate the incidence of occurrence for 29 different sightings around Africa between 1829 and 1949, compared to 51 British Isles (BI) sightings between 1809 and 2000, 12 different Nova Scotian (NS) sightings between 1787 and 1939, 20 different sightings in New England (NE) between 1831 and 1892, and 28 different sightings in the Western Pacific (WP) between 1854 and 1962 (see France 2020 and other sources therein).
| Attribute | Africa | BI | NS | NE | WP |
|---|---|---|---|---|---|
| Notable length | 20 | 21 | 11 | 9 | 20 |
| Body composed of a series of irregular, jointed component parts (multiple humps, coils, or a ridge) | 13 | 24 | 7 | 9 | 15 |
| Rapid speed of movement | 7 | 10 | 5 | 3 | 4 |
| Notable flexibility of body | 1 | 4 | 3 | 1 | 1 |
| Vertical undulating movement of body segments | 6 | 24 | 5 | 3 | 10 |
| Body components likened to kegs or barrels, and sometimes scales or saucers | 8 | 2 | 8 | 2 | 6 |
| Oblivious of surroundings or impervious to disturbance | 3 | 5 | 4 | 2 | 3 |
| Overall body likened to a string of floats, kegs or buoys | 3 | 0 | 3 | 3 | 0 |
| Obvious trailing wake or water disturbance | 6 | 6 | 6 | 2 | 6 |
| Floating motionless, gently swaying in waves, or moving very slowly | 8 | 5 | 1 | 2 | 6 |
| Narrow, tapering, sinuous, snake/eel- 16 like shape, often with absence of a caudal fin or lateral appendages | 16 | 14 | 8 | 3 | 15 |
| Extended body pulled down into water, thrown up into the air, or thrashing about on the surface | 4 | 5 | 0 | 1 | 3 |
| Presence of a horn, spike, spine, mane or other protuberance (sometimes identified as a head) | 15 | 11 | 3 | 2 | 9 |
As the case for UMOs from North America, the Western Pacific and the British Isles, those observed in African waters are also described as having body components that imply entangled debris. Several eyewitnesses liken bodies to casks and refer to the presence of scales. As previously mentioned, wooden barrels and glass-balls were used as floats on fishing nets throughout the nineteenth century, which can be advanced to parsimoniously explain the present observations. And as with North American UMOs (Table 4), several anecdotes about African ‘sea serpents’ explicitly describe their overall bodies as resembling a rope length of fishing-net floats. This is the closest the evidence comes to a ‘smoking gun’ on a par with that observed for the Gloucester UMO, which was almost certainly an entangled animal.
Other telling evidence supporting the contention that these African UMOs, believed to have been mysterious creatures were merely mundane animals who had the misfortune of becoming entangled can be discerned through parsing descriptions of the physical attributes of what were described as body ‘coils’ and ‘hair.’ It is biomechanically impossible for the body of an animal flexible enough to engage in undulating movement to be composed of a set of arches that are elevated above the surface of the water. Furthermore, the remarkable fact that body segments described as being composed of a series of ‘knobs protruding above the water’ maintained their form as ‘solid and static’ even when the animal was at rest, proves unequivocally that the components could not be biological. Added to this are anecdotes describing long, brown filaments believed to have been hair draped or ‘washed about the back’ of UMOs, in several cases being likened to attached mats of kelp. This, of course, is just how animals look when entangled in fishing nets, as can be seen in many online photos today. As was the case for the entangled Gloucester UMO, the presence of horns or spars elevated above the water for several African UMOs connote either marlinspikes and fids – instruments used to intertwine lengths of hemp ropes – or marker poles attached to barrel floats set on the ends of deployed fishing nets.
The frequent mention made of the rapid motion of African UMOs is consistent with observations from other regions (Table 4), and provides confirmatory evidence that what was being observed must have had a non-biological explanation. For, in the absence of observable large pectoral fins or a bilobate caudal tail, it is of course impossible for a serpentine animal relying upon only vertical undulations to generate sufficient pressure against the water to achieve such speed (van Weerden et al. 2013). Obviously the propulsive force must come from elsewhere. A plausible explanation is that it comes from swimming animals that are either temporarily or permanently invisible beneath the surface while they pull trains of buoyant entangled material. Furthermore, marine animals of noted mobility have body shapes that have specifically evolved to limit as much drag as possible. That African UMOs are described as generating considerable water disturbance when swimming, once again points to a non-biological cause.
The caterpillar-like, up-and-down motion of African UMOs is counter to the horizontal flexure on which all fish and sea snakes rely for propulsion. Cryptozoologists, when they acknowledge this inconvenient truth, use it to go on to justify their claims that, as only sea mammals can bend their bodies in the vertical plane, ipso facto, it means that sea serpents must be relic species of ancient whales or seals that have somehow escaped both extinction and discovery. Alternatively, the parsimonious explanation, referred to as being ‘the best explanation yet’ (Deedy 2017), is that the undulations are merely a string of fishing-net floats bobbing up and down on the surface of the water.
It is much easier to reach a conclusion that sightings of many-humped ‘sea serpents’ represent evidence for the early, pre-plastic entanglement of marine fauna than it is to identify the encumbered animals at the front-end of debris chains. As the case for other locations (Table 4), the segmented bodies of African UMOs were often spotted resting motionless upon the surface, sometimes followed by their sudden submergence for prolonged periods. This suggests that, for these particular cases, the entangled animals were likely to have been large fish. Similar to UMOs from elsewhere (Table 4), those from Africa were also inattentive to their surroundings. This included, for example, being dragged beneath a ship, as well as being unresponsive to physical assault. It is impossible to avoid the conclusion that these observed ‘creatures’ could only have been inanimate objects.
African UMOs were occasionally observed violently thrashing about on the surface. The cryptozoological literature contains frequent encounters described in a similar fashion, which are interpreted therein as recognised animals engaged in battle with imagined sea serpents. More likely (see illustrations and descriptions in France 2016, 2017), however, and in agreement with dramatic descriptions of modern-day animals struggling to free themselves from fishing gear (e.g. Johnson 2005), these encounters almost certainly represent some of the earliest observations made of the susceptibility of marine life to entanglement.
Africa and the Anthropocene
The present work, supporting similar historical investigations undertaken for other regions (France 2020 and references therein), strongly suggests the global ubiquity of early, pre-plastic entanglement. Of interest is that nineteenth-century artisanal fisheries in Africa, just like those in the Western Pacific, both exhibit the same phenomena, and actually do so in proportions (in relation to all documented UMO sightings for their respective regions) that are higher than those similarly documented contemporaneously for other, more developed locations such as North America and Europe, whose historical fisheries have been extensively studied. This implies that it is not only the famous ‘charismatic terrestrial megafauna’ of Africa that have borne the impact of humans for a prolonged period, but its marine fauna as well. Hecht’s (2018) contention of an ‘African Anthropocene’ therefore seems apropos.
The timing of what is believed to constitute the beginning of the Anthropocene is a topic of lively debate (e.g. Biello 2015), much of it originating from mentalities ensconced within disciplinary silos. This has led some to proclaim that there are in fact many different anthropocenes, rather than a singular Anthropocene (Edgeworth et al. 2015; Ruddiman et al. 2015). What is clear is that, from a perspective of biodiversity and species extinctions (Kolbert 2015), that this particular anthropocene had its start as a slow drum roll rather than as a sudden bang, given that it was initiated far in advance of the detonation of atomic bombs, the onset of the industrialisation, or even the Neolithic Revolution (e.g. Wong 2012). And it is for this reason that, of all scholars documenting anthropogenic change, it is wildlife biologists who are most susceptible to feeling bereft, something recognised more than half a century ago by Aldo Leopold in his poignant statement about feeling increasingly alone ‘in a world of wounds.’
That today’s oceans harbour but a vestige of their original bounty is a sad reality (Mowat 1997). Much of the work of marine environmental historians (e.g. Rick and Erlandson 2008; Kittinger et al. 2015) has been concerned with documenting the widespread damage to targeted fish stocks brought about by non-sustainable harvesting practices. The present environmental history, building upon previous research for other regions, suggests that animals have also been incidental victims of deployed fishing equipment for much longer than is customarily believed. This collateral damage caused by normal fishery operations – entanglement – should therefore be considered part and parcel of comprehensive assessments made of the history of anthropogenic stress experienced by marine megafauna, animals whose sizes once led them to be perceived as ‘sea monsters’ (Mazzoldi et al. 2019).
What this means is that there is no sharp discontinuity between old and new technology. The fishery narrative is therefore not one of an onslaught of a destructive modernity contrasted to a halcyon pastoralism; in others words, bad plastic verses good natural fibres. In terms of anthropogenic stress to wildlife, there is no sudden arrival of a ‘machine in the garden.’ The use of new technology for resource extraction merely extends and amplifies the older techniques. With its history of fishing extending for thousands of years (Fagan 2017), Africa has always been an active player in the Anthropocene, or at least in the particular, small ‘a’ anthropocene concerned with biodiversity. We know, for example, that two-thirds of the continent’s large carnivores have been hunted to extinction (see Biello 2013). In the absence of widespread, quantitative catch records of the type customarily examined by environmental historians in Europe and North America, the present anecdotal evidence (in the sense of Pauly 1995), vis-à-vis inferred entanglement in fishing/hunting gear or other maritime debris, suggests that it is likely Africa’s marine fauna have also been impacted for a considerable time. In this light, and irrespective of either uranium (Hecht 2018) or plastic, it seems the continent has not for some millenia been outside of what can be considered to be modernity and its unwelcome ramifications.
Monsters, Real and Metaphorical, of the Anthropocene
Because sea serpents and lake monsters are mental constructs (Meurger and Gagnon 1988), they are always there swimming about just beneath the surface of our collective consciousness, ready to emerge at the slightest provocation, often coinciding with periods of societal anxiety.3 For example, a climate of uncertainty during the Victorian Age, a time when sea monsters figured prominently (Barber 1980), was fuelled by the promulgation and widespread discussion of competing theories and beliefs. Lyons (2009) suggests the period be better known as the ‘age of contradictions’ or ‘age of transitions’ than the customary ‘age of science.’ To Victorians, sea monsters embodied alternative modes of understanding the natural world (Ritvo 1998) because they challenged Linnaean classification. If composite creatures such as these defy categorisation, so the reasoning goes, they provoke a ‘crisis’ (Cohen 2020). Their existence undermines our place of wisdom at the apex of Creation, a position already shaken by Darwin’s (r)evolutionary theory (Lyons 2009; France 2017). Many Victorians were also apprehensive about the precipitous advent of modernity brought about by industrialisation (Blake’s ‘Satanic mills’), and what that meant in terms of a discontinuity from the world beforehand, something in which sea monsters figured (Lyons 2009). In Jules Verne’s 1870 novel Twenty Thousand Leagues Under the Sea, for example, an expedition sets out in search of a fearful sea monster which of course turns out to be an equally frightening technologically advanced submarine piloted by a mysterious figure (Captain Nemo – whose name in Latin means ‘nobody’) engaging in monstrous behaviour (France 2017). But the most significant role that sea monsters played in terms of societal anxiety in the nineteenth century concerned the inconvenient truth their very existence represented.
Paleontological discoveries of ancient ‘sea dragons’ throughout the nineteenth century had a profound effect upon Victorian society by giving scientific legitimacy to the idea that living fossils in the form of sea monsters might still be swimming about in incompletely explored oceans (Lyons 2009; Paxton and Naish 2019). Theology’s controversy with fossils in the nineteenth century concerned the concept of deep time and the eschatological anxiety of ‘temporal vertigo’ (see McGowan-Hartman 2013) that ensued when the hitherto cosy view of six thousand years of world history was suddenly extended to millions of years. To the religious minded, the idea that fossils represented extinct animals was unfathomable since it made no sense for God to create animals that disappeared aeons before humans arrived on the scene. Ergo, ancient sea dragons in the form of their descendants (‘sea monsters’) must by necessity still exist out there somewhere (France 2019a). Rather than invoking terror, sightings of sea monsters such as those described here, brought a form of epistemological comfort.
Fast forward to the present, when many anomalous or preternatural monsters can be logically explained by wildlife biologists, medical experts and geomythologists, and a completely different situation exists. So whereas to the Victorians monsters were regarded as being very much real corporeal beings, monsters today, because they have always represented our worst fears (Asma 2009), have come to be looked upon as convenient metaphors of social-ecological disorder in the Anthropocene (e.g. Tsing et al. 2017; Giulani 2020) (perhaps no more overtly so than in the dozens of sea monster films produced during the 1950s fuelled by concerns about the threat of atomic radiation). This is part of the long tradition of monsters presaging turbulent times (Asma 2009), and makes perfect sense since the word ‘monster’ derives from the Latin word monstrum, which itself denotes warning; i.e. as signs, they ‘demonstrate and portend the future’ (italics in original, Neville 2001: 107). And of course there is no more famous monster-as-a-metaphor in this regard than the creature brought to life in Mary Shelley’s Frankenstein.
Shifting social constructions influence perceptions of monstrosity. In the modern era, the term is used less to apply to physical appearance than to horrible behaviour (Weinstock 2016), the latter often as much in relation to treatment to, rather than acts by, the titular ‘monster’. Long thought to be a cautionary tale about scientific hubris, Frankenstein has been repeatedly reimagined (Baldick 1987; Six and Thompson 2016). Wood (2016) draws a parallel between the hordes of wandering refugees displaced by the eruption of Tambora in 1815 and the ‘Frankenstein weather’ that followed. In New England, the ensuing famous ‘year within a summer’ triggered a sequence of adaptations which led, in extremis, to a resource shift to mackerel (Alexander et al. 2017). The result was the sudden and massive deployment of fishing gear in offshore waters, which in turn caused the nonlethal entanglement of an animal and the consequent folkloric invention of what became known as the Gloucester Sea Serpent (France 2019b).
As Olsen and Olsen (2001: 6) state: ‘monsters are, in effect, the tangible representations of this past or present otherness.’ In addition to being emblematic of freakish nature, monsters also function to ‘explore both the good and bad limits of human potential’ (Olsen and Olsen 2001: 12). Because of what it represents in terms of ‘ecobiocentric ideals’ (Hammond 2004) and expressions of inhumanity, Dr Frankenstein’s Creature is coming to be regarded as the perfect metaphor for the Anthropocene (MacCormack 2018). Therefore, if Higgins (2016) is correct about ‘Why a Volcano, Frankenstein, and the Summer of 1816 are Relevant to the Anthropocene,’ wherein the walking undead of Shelley’s monster symbolises inhumane living conditions and consequent displacement due to either climate change or conflict (Marshall 2020), then the swimming undead of sea monsters – ‘creatures’ entangled in fishing gear, whether synthetic material or natural fibres, likewise provides a powerful metaphor for the destruction of marine ecosystems during the Anthropocene. For, as Higgins (2016) wrote, modern monstrosity, considered in this regard, ‘is not so much concerned with what we “do” to nature or vice versa, but with the entanglement of human and nonhuman agents’ (my italics). And in this we have – to use an appropriately serpentine symbol – an ouroboros situation, wherein sea monsters in Western culture at one time were frequently used as metaphors (e.g. the Biblical Leviathan, moralising medieval bestiaries, Renaissance map illustrations, and the rhetoric of colonisation), then in the late-modern period when they came to be regarded as real creatures both new to science and providing comfort to threatened religious dogma, to now, in the troubling times of our zombie-obsessed twenty-first century (Korovkin and Stephenson 2010; Stratton 2020), wherein once again sea monsters may serve as a metaphor for the ‘dissevering’ (to quote the poet Robinson Jeffers in France 1992) of human-nature ‘integrity’.
Notes
- It is important to emphasise that the focus of this research is on entanglement in a literal or physical sense, not as a notional metaphor, such as, for example, used by some Anthropocene scholars (e.g. Holm & Taffel 2016; Hamilton 2017). Nor are we concerned here with the general conceptual rubric by which anthropologists have come to regard multispecies interactions or, in their parlance, ‘entanglements’ (e.g. Carrithers et al. 2011; Giraud 2019); i.e. as a form of new wine in old bottles, following as it does in the long established tradition of environmental geographers, deep ecologists, ethnobiologists and eco-theologians who have long considered humans to share the fates of other species in conjoined social-ecological systems rather than existing as autonomous beings. Likewise, the whole troubling issue of widespread oceanic pollution by microplastics is not germane to the present investigation. ⮭
- Such thinking persisted throughout the Enlightenment and well into the late modern period. After all, gorillas – scientifically known from only the middle of the nineteenth century (Newman 2017) – which derive their name from Hellenic Greek meaning ‘tribe of hairy women,’ represented an unsettling uncertainty in debates about natural science and human evolution (Scott 2020). That Africa was inhabited by many such strange creatures was long the accepted norm (van Druzer 2016): Pliny the Elder remarked that the continent was always yielding new phenomena; Herodotus mentioned that its western edge is where ‘huge snakes are found… and a great many other creatures of by no means a fabulous kind’ (396); such that it is no surprise that various medieval mappa mundi depicted monsters inhabiting the southern edge of the continent. In more recent times, Edgar Rice Burroughs, in Tarzan the Terrible, had his eponymous hero encounter a valley filled with dinosaurs in Africa. And because prehistoric (or paleo- or crypto-) fiction serves as the wellspring for much of cryptozoological fancy (Mullis 2019), there are even those today whom maintain that real dinosaurs, regarded as folkloric monsters by those whom they consider to be simplistic locals, continue to inhabit Africa lakes and wetlands (Makal 1987; Nugent 1993; see Loxton and Prothero 2015 for a scathing synopsis of this modern-day picaresque-cum-Pythonesque charade of anachronistic natural history exploration). Let us not forget that the favourite animal touted by cryptozoologists to justify their claims that the seas are filled with prehistoric animals, the coelacanth (Latimeria chalumnae), was first collected off the east coast of Africa. All this is in addition to an indigenous folklore filled with a panoply of imaginary creatures (e.g. Walsh and Goldman 2020), including mysterious water beings, some of whose origins lie in a cross-cultural fertilisation of African and European beliefs (Drewal 2016). It is easy to see why Saunders (2018) stated that ‘Africa’s blank spaces on the map have long been filled with monstrous creatures that fuel the western imagination,’ and pointed to a history of Africa being used ‘as a palimpsest to construct fantastical tales.’ Therefore, the perception of a ‘Dark Continent’ monstrosity held by Victorian Europeans remains appealing, even today, as witness to a recent film entitled Monsters: Dark Continent and a newspaper article with the headline ‘Are Scions of the Prehistoric Monsters That Once Roamed the Earth Still to Be Found Within Little-Known Parts of the Dark Continent?’ ⮭
- Monster theory has itself emerged as a fascinating and vibrant field of scholarship (e.g. Mittman and Dendle 2016; Weinstock 2020). It is impossible, in the limited space available, to explore how leading theorists such as Latour, Shellenberer and Haraway have grappled with the intriguing topic of the complex relationships that exist between our twin progeny: monsters and technology. As Cohen (2020) explains, monsters are our children, asking us to question our perception of the world and why we have created them in our imagination. Instead, and mimicking the strategy of Sarah Perry in her award-winning 2016 novel about Victorian concepts of science and superstition, The Essex Serpent (which interestingly, given the gist of the present thesis, was published by a company called ‘Serpent’s Tail’), I have used the idea of sea monsters more as a convenient leitmotif throughout, while recognising that I am but skirting the edge of the vast sea of monster theory. But there is one interesting concept that should be mentioned, if only so cursorily, and that concerns the idea of sea monsters as composite or chimera creatures. Many sightings of sea monsters follow the same general trajectory: the Victorian observers first spot an animal they immediately recognise – a whale or a turtle or a seal – but then they almost immediately notice the long extended ‘tail’ which causes them to alter their initial identification, leading to the conclusion that the UMO defies the norm and must, therefore, ipso facto, be a sea monster. In doing so, they are simply adhering to the zeitgeist wherein such creatures are born from the confluence of Biblical allegory, classical antiquity, Nordic mythology, and recent paleontological discoveries of the era (Loxton and Prothero 2015; France 2019a). Hybrid creatures have always played a prominent role in discussions of natural history (e.g. Brink-Roby 2008; Ritvo 2010), with sea monsters becoming the locus for several boundary disputes in the culture of Victorian science (Lyons 2009). The present illation that many so-called sea monsters were in fact known animals that had become entangled in anthropogenic material extends this discussion to issues of the Anthropocene because it indicates that the creatures are in a sense natural-technological fusions, or cyborgs. In this way, such ‘monsters’ represent a corporeal counterpoint to the vast scholarship that frequently regards such constructs in a metaphorical sense (e.g. Graham 2003). Clearly this is an area of monster theory worthy of future exploration/elaboration. ⮭
Competing Interests
The author has no competing interests to declare.
References
1 Aiken, W. R. G., & Purser, J. (1936). The Preservation of Fibre Ropes for Use in Sea-Water. Plymouth Laboratory New Series, 20, 643–654. DOI: http://doi.org/10.1017/S0025315400058227
2 Akyeampong, E., Bates, R. H., Nunn, N., & Robinson, J. A. (2014). Africa’s Development in Historical Perspective. Cambridge: Cambridge University Press. DOI: http://doi.org/10.1017/CBO9781139644594
3 Al-Abdulrazzak, D., Naidoo, R., Palomares, M. L. D., & Pauly, D. (2012). Gaining Perspective on What We’ve Lost: The Reliability of Encoded Anecdotes in Historical Ecology. PLOS One, 7(8), 1–5. DOI: http://doi.org/10.1371/journal.pone.0043386
4 Alexander, K. E., et al. (2017). Tambora and the Mackerel Year: Phenology and Fisheries During an Extreme Climate Event. Science Advances, 3(1), 1–18. DOI: http://doi.org/10.1126/sciadv.1601635
5 Alverson, D. L., Freeberg, M. H., Murawski, S. A., & Pope, J. G. (1994). A Global Assessment of Fisheries Bycatch and Discards. Fisheries and Oceans Fisheries Technical Paper, 339.
6 Anderson, F. (2016). The Dark Continent? Images of Africa in European Narratives about the Congo. Aarhus: Aarhus University Press.
7 Anonymous. (2019). Whale washed up in Caithness tangled in Canadian fishing gear. BBC News, 03/06/19. www.bbc.com/news/uk-scotland-highlands-islands-48497046.
8 Asma, S. T. (2009). On Monsters: An Unnatural History of Our Worst Fears. Oxford: Oxford University Press.
9 Baldick, C. (1987). In Frankenstein’s Shadow: Myth, Monstrosity, and Nineteenth-century Writing. Clarendon Press.
10 Barber, L. (1980). The Heyday of Natural History, 1820–1870. New York: Doubleday & Company.
11 Beinart, W. (2000). African History and Environmental History. African Affairs, 99(395), 269–302. DOI: http://doi.org/10.1093/afraf/99.395.269
12 Bekker-Nielsen, T., & Casola, D. B. (Eds.) (2010). Ancient Nets and Fishing Gear. Aarhus: Aarthus University Press.
13 Belozerskaya, M. (2009). The Medici Giraffe: and Other Tales of Exotic Animals and Power. Boston, MA: Little, Brown and Company.
14 Biello, D. (2015). Did the Anthropocene Begin in 1950 or 50,000 Years Ago? Scientific American, 2 April.
15 Brantlinger, P. (1985). Victorians and Africans: The Genealogy of the Myth of the Dark Continent. Critical Inquiry, 12(1), 166–203. DOI: http://doi.org/10.1086/448326
16 Brewer, D. J., & Friedman, R. F. (1989). Fish and Fishing in Ancient Egypt. Cairo: The American University in Cairo Press.
17 Brink-Roby, H. (2008). Siren Canora: The Mermaid and the Mythical in Late Nineteenth-Century Science. Archives of Natural History, 35(1), 1–14. DOI: http://doi.org/10.3366/E0260954108000041
18 Brown, C. M. (1990). A Natural History of the Gloucester Sea Serpent: Knowledge, Power, and the Culture of Science in Antebellum America. American Quarterly, 42(3), 402–436. DOI: http://doi.org/10.2307/2712941
19 Carrithers, M., Bracken, L. J., & Emery, S. (2011). Can a Species be a Person? A Trope and its Entanglements in the Anthropocene Era. Current Anthropology, 52(5), 661–685. DOI: http://doi.org/10.1086/661287
20 Carruthers, J. (2004). Africa: Histories, Ecologies and Societies. Environment and History, 10(4), 379–406. DOI: http://doi.org/10.3197/0967340042772649
21 Charmaz, K. (2007). Constructing Grounded Theory: A Practical Guide Through Qualitative Analysis. Thousand Oaks, CA: Sage Publications.
22 Cliff, G., Dudley, S., Ryan, P. G., & Singleton, N. (2002). Large Sharks and Plastic Debris in KwaZulu-Natal, South Africa. Marine and Freshwater Research, 53, 575–581. DOI: http://doi.org/10.1071/MF01146
23 Cohen, J. J. (2020). Monster Culture (Seven Theses). In J. A. Weinstock (Ed.). The Monster Theory Reader (pp. 37–57). Minneapolis, MN: University of Minnesota Press. DOI: http://doi.org/10.5749/j.ctvtv937f.5
24 Das, N. (2009). Occam’s Razor and Cryptozoology. The Scienta Review. MIT.
25 Dearing, J. A., et al. (2015). Social-ecological Systems in the Anthropocene: The Need for Integrating Social and Biophysical Records at Regional Scales. The Anthropocene Review, 2(3), 220–246. DOI: http://doi.org/10.1177/2053019615579128
26 de Camp, L. S., & Crook de Camp, C. (1985). The Day of the Dinosaur. New York: Bonanza Books.
27 Deedy, A. (2017). Hark! A sea monster! (Oh, no, just a dying whale). All those strange sea monster sightings in days of yore? This may be the best explanation yet. Hakai Magazine 8 February. Available at: https://www.hakaimagazine.com/news/hark-sea-monster-oh-no-just-dying-whale
28 Derraik, J. G. B. (2002). The Pollution of the Marine Environment by Plastic Debris: A Review. Marine Pollution Bulletin, 44(9), 842–852. DOI: http://doi.org/10.1016/S0025-326X(02)00220-5
29 Diamond, J. (2007) Guns, Germs and Steel: The Fate of Human Societies. New York: W. Norton.
30 Drewal, H. J. (2016). Beauteous Beast: The Water Deity Mami Wata in Africa. In A. S. Mittman & P. J. Dendle (Eds.), The Ashgate Research Companion to Monsters and the Monstrous (pp 77–102). New York: Routledge.
31 Edgeworth, M., deB Richter, D., Waters, C., Haff, P., Neal, C., & Price, S. J. (2015). Diachronous Beginnings of the Anthropocene: The Lower Boundary Surface of Anthropogenic Deposits. The Anthropocene Review, 2(1), 33–58. DOI: http://doi.org/10.1177/2053019614565394
32 Ellis, E., Maslin, M. N., Boivin, N., & Bauer, A. (2016). Involving Social Scientists in Defining the Anthropocene. Nature, 540(7632), 192–193. DOI: http://doi.org/10.1038/540192a
33 Fagan, B. (2017). Fishing: How the Sea Fed Civilization. New Haven, CN: Yale University Press. DOI: http://doi.org/10.2307/j.ctv1bvnff7
34 Fama, E. (2012). Debunking a Great New England Sea Serpent. Tor.com, 16 August. Available at: https://www.tor.com/2012/08/16/debunking-a-great-new-england-sea-serpentTor.com
35 France, R. L. (1992). Gaian Integrity: A Clarion Precept for Global Preservation. The Trumpeter, 4, 159–163.
36 France, R. L. (2016). Reinterpreting Nineteenth-century Accounts of Whales Battling ‘Sea Serpents’ as an Illation of Early Entanglement in Pre-Plastic Fishing Gear or Maritime Debris. International Journal of Maritime History, 28(4), 686–714. DOI: http://doi.org/10.1177/0843871416667434
37 France, R. L. (2017). Imaginary Sea Monsters and Real Environmental Threats: Reconsidering the Famous Osborne, ‘Moha-moha’. Valhalla, and ‘Soay Beast’ Sightings of Unidentified Marine Objects. International Review of Environmental History, 3(1), 63–100. DOI: http://doi.org/10.22459/IREH.03.01.2017.07
38 France, R. L. (2018). Illustration of an 1857 ‘Sea Serpent’ Sighting Reinterpreted as an Early Depiction of Cetacean Entanglement In Maritime Debris. Archives of Natural History, 45(1), 111–117. DOI: http://doi.org/10.3366/anh.2018.0486
39 France, R. L. (2019a). Disentangled: Ethnozoology and Environmental Explanation of the Gloucester Sea Serpent. Wageningen, NL: Wageningen Academic Press. DOI: http://doi.org/10.3920/978-90-8686-886-5
40 France, R. L. (2019b). Extreme Climatic Upheaval, Emergency Resource Adaptation, and the Emergence of Folkloric Belief: Geomythic Origin of Sea Serpents from Animals Becoming Entangled in Fishing Gear During New England’s Nineteenth-Century Social-Ecological Crisis. Human Ecology, 47, 499–513. DOI: http://doi.org/10.1007/s10745-019-00097-5
41 France, R. L. (2020). Historical Anecdotes of Fishing Pressure: Misconstrued ‘Sea Serpent’ Sightings Provide Evidence For Antecedent Entanglement of Marine Biota in the British Isles. Fish and Fisheries online. DOI: http://doi.org/10.1111/faf.12505
42 France, R. L. (In press). Ethnozoology of Egede’s ‘Most Dreadful Monster’, the Foundational Sea Serpent. Contributions in Ethnobiology, Contributions in Ethnobiology monograph series, Denton, TX: Society of Ethnobiology.
43 France, R., & Rigg, C. (1998). Survey of the Global Distribution of Biodiversity Research Published in Five International Journals: Is There a Tropical Bias? Tropical Biodiversity, 5, 161–166.
44 Freedman, J. A., & Noakes, D. L. G. (2002). Why Are There No Really Big Fishes? A Point-of-view on Maximum Body Size in Teleosts and Elasmobranchs. Review in Fish Biology and Fisheries, 12, 403–416. DOI: http://doi.org/10.1023/A:1025365210414
45 Galbreath, G. J. 2015. The 1848 ‘Enormous Serpent’ of the Daedalus Identified. The Skeptical Inquirer, 39(5), 42–46.
46 Gardner, D. (2007). Whale Survives Harpoon Attack 130 years ago to Become ‘World’s Oldest Mammal’. The Daily Mail, 13 June.
47 Giraud, E. H. (2019). What Comes After Entanglement? Activism, Anthropocentrism, and an Ethics of Exclusion. Durham, NC: Duke University Press. DOI: http://doi.org/10.1515/9781478007159
48 Giuliani, G. (2020). Monsters, Catastrophes and the Anthropocene: A Postcolonial Critique. London: Routledge. DOI: http://doi.org/10.4324/9781351064866
49 Goode, G. B., et al. (1884). The Fisheries and Fishery Industries of the United States. National Oceanographic and Atmospheric Administration reprint. (Online access, numerous sources).
50 Graham, E. L. (2003). Frankenstein and Cyborgs: Visions of The Global Future in an Age of Technology. Studies in Christian Ethics, 16(1), 29–43. DOI: http://doi.org/10.1177/095394680301600103
51 Gregory, M. R. (2009). Environmental Implications of Plastic Debris in Marine Settings – Entanglement, Ingestion, Smothering, Hangers-On, Hitch-Hiking and Alien Invasions. Philosophical Transactions of the Royal Society B, 364(1526), 2013–2025. DOI: http://doi.org/10.1098/rstb.2008.0265
52 Grigson, C. (2018). Menagerie: The History of Exotic Animals in England. Oxford: Oxford University Press.
53 Hamilton, S. (2017). Securing Ourselves From Ourselves? The Paradox of ‘Entanglement’ in the Anthropocene. Crime, Law and Social Change, 68, 579–595. DOI: http://doi.org/10.1007/s10611-017-9704-4
54 Hammond, K. (2004). Monsters of Modernity: Frankenstein and Modern Environmentalism. Cultural Geographies, 11(2), 181–198. DOI: http://doi.org/10.1191/14744744004eu301oa
55 Hecht, G. (2018). Interscalar Vehicles for an African Anthropocene: On Waste, Temporality, and Violence. Cultural Anthropology, 33(1), 109–1412. DOI: http://doi.org/10.14506/ca33.1.05
56 Herre, A. W. (1942). The Whale Shark in the Philippines. The Scientific Monthly, 55(2), 151–158.
57 Heuvelmans, B. (1968). In the Wake of the Sea-Serpents. New York: Hill and Wang.
58 Higgins, D. (2016). Why a volcano, Frankenstein, and the summer of 1816 are relevant to the Anthropocene. The Conversation, 12 September. Available at https://theconversation.com/why-a-volcano-frankenstein-and-the-summer-of-1816-are-relevant-to-the-anthropocene-64984.
59 Holm, N., & Taffel, S. (2016). Ecological Entanglements in the Anthropocene. Washington, DC: Lexington Books.
60 Hsieh, H.-F., & Shannon, S. E. (2005). Three Approaches to Qualitative Content Analysis. Qualitative Health Research, 15(9), 1277–1288. DOI: http://doi.org/10.1177/1049732305276687
61 Jackson, J. B. C., Alexander, K. E., & Sala, E. (2011). Shifting Baselines: The Past and the Future of Ocean Fisheries. Washington, DC: Island Press. DOI: http://doi.org/10.5822/978-1-61091-029-3
62 Jarosz, L. (1992). Constructing the Dark Continent: Metaphor as Geographic Representation of Africa. Geografiska Annaler: Series B, Human Geography, 74(2), 105–15. DOI: http://doi.org/10.1080/04353684.1992.11879634
63 Johnson, T. (2005). Entanglements: The Intertwined Fates of Whales and Fishermen. Gainesville, FL: University Press of Florida.
64 Kaplan, S. (2016). By 2050, there will be more plastic in the world’s oceans than fish, study says. The Washington Post, 2016/01/20.
65 Kittinger, J. N., McClenachan, L., Gedan, K. B., & Blight, L. K. (Eds.) (2015). Marine Historical Ecology in Conservation: Applying the Past to Manage for the Future. Berkeley, CA: University of California Press. DOI: http://doi.org/10.1525/california/9780520276949.001.0001
66 Kolbert, E. (2015). The Sixth Extinction: An Unnatural History. London: Picador.
67 Korovkin, M., & Stephenson, P. (2010). Zombie Factory: Culture, Stress & Sudden Death. London: Green Frigate Books (Libri Publishing).
68 Laist, D. W. (1997). Impacts of Marine Debris: Entanglement of Marine Life in Marine Debris Including a Comprehensive List of Species with Entanglement and Ingestion Records. In J. M. Coe & D. B. Rogers (Eds.), Marine Debris: Sources, Impacts and Solutions (pp. 56–79). New York: Springer. DOI: http://doi.org/10.1007/978-1-4613-8486-1_10
69 Lehn, W. H., & Schroeder, I. I. (2004). The Hafstramb and Margygr of the King’s Mirror: An Analysis. Polar Record, 40(2), 121–134. DOI: http://doi.org/10.1017/S0032247403003255
70 Lloyd, J. B. (1971). African Animals in Renaissance Literature and Art. Oxford: Clarendon Press.
71 Loxton, D., & Prothero, D. R. (2015). Abominable Science! Yeti, Nessie, and Other Famous Cryptids. New York: Columbia University Press.
72 Lyons, S. L. (2009). Species, Serpents, Spirits, and Skulls: Science at the Margins in the Victorian Age. Albany, NY: State University of New York Press.
73 MacCormack, P. (2018). Two centuries on, Frankenstein is the perfect metaphor for the Anthropocene Era. The Conversation, 1 January. Available at https://theconversation.com/two-centuries-on-frankenstein-is-the-perfect-metaphor-for-the-anthropocene-era-88679
74 Makal, R. (1987). A Living Dinosaur? In Search of Mokele-Mbembe. Leiden: Brill.
75 Marshall, A. (2020). Did a volcanic eruption in Indonesia really lead to the creation of Frankenstein? The Conversation, 23 January.
76 Mazzoldi, C., et al. (2019). From Sea Monsters to Charismatic Megafauna: Changes in Perception and Use of Large Marine Animals. PLOS One, 31 December. DOI: http://doi.org/10.1371/journal.pone.0226810
77 McCaskill, J. (2009). Conserving Waterlogged Rope: A Review of Traditional Methods and Experimental Research with Polyethylene Glycol. (Masters Thesis, College Station, Texas A&M University).
78 McClain, C. R., et al. (2015). Sizing Ocean Giants: Patterns of Intraspecific Size Variation in Marine Megafauna. PeerJ3: e715. DOI: http://doi.org/10.7717/peerj.715
79 McClenachan, L., Ferretti, F., & Baum, J. K. (2012). From Archives to Conservation: Why Historical Data are Needed to Set Baselines for Marine Animals and Ecosystems. Conservations Letters, 5(2), 349–359. DOI: http://doi.org/10.1111/j.1755-263X.2012.00253.x
80 McGowan-Hartman, J. (2013). Shadow of the Dragon: The Convergence of Myth and Science in Nineteenth Century Paleontological Imagery. Journal of Social History, 47(1), 47–70. DOI: http://doi.org/10.1093/jsh/sht048
81 Meine, C. (1999). It’s About Time: Conservation Biology and History. Conservation Biology, 13(1), 1–3. DOI: http://doi.org/10.1046/j.1523-1739.1999.013001001.x
82 Melles, S. J., et al. (2019). Diversity of Practitioners Publishing in Five Leading International Journals of Applied Ecology and Conservation Biology, 1987–2015, Relative to Global Biodiversity Hotspots. Ecoscience, 26(4), 323–340. DOI: http://doi.org/10.1080/11956860.2019.1645565
83 Meurger, M., & Gagnon, C. (1988). Lake Monster Traditions: A Cross-Cultural Analysis. London: Fortean Tomes.
84 Meyer, M. A., Best, P. B., Anderson-Reade, M. D., Cliff, G., Dudley, S. F. J., & Kirkman, S. P. (2011). Trends and Interventions in Large Whale Entanglement along the South African Coast. African Journal of Marine Science, 33(3), 429–439. DOI: http://doi.org/10.2989/1814232X.2011.619064
85 Miller, J. (2012). Empire and the Animal Body: Violence, Identity and Ecology in Victorian Adventure Fiction. London: Anthem Press. DOI: http://doi.org/10.7135/UPO9780857285492
86 Mittman, A. S., & Dendle, P. J. (Eds.). (2016). The Ashgate Research Companion to Monsters and the Monstrous. New York: Routledge. DOI: http://doi.org/10.4324/9781315241197
87 Mowat, F. (1997). Sea of Slaughter. London: McClelland-Bantam.
88 Mullis, J. (2019). Cryptofiction! Science Fiction and the Rise of Cryptozoology. In D. Caterine & J. W. Morehead (Eds.), The Paranormal and Popular Culture: A Postmodern Religious Landscape (pp. 240–252). Abingdon: Routledge. DOI: http://doi.org/10.4324/9781315184661-19
89 National Oceanic and Atmospheric Administration. [NOAA]. (2014). Report on the Entanglement of Marine Species in Marine Debris with an Emphasis on Species in the United States. Silver Spring, MD: National Oceanic and Atmospheric Administration Marine Debris Program.
90 Neilson, J. L., Straley, J. M., Gabriele, C. M., & Hills, S. (2009). Non-lethal Entanglement of Humpback Whales (Megaptera novaeangliae) in Fishing Gear in Northern Southeast Alaska. Journal of Biogeography, 36(3), 452–464. DOI: http://doi.org/10.1111/j.1365-2699.2007.01820.x
91 Neville, J. (2001). Monsters and Criminals: Defining Humanity in Old English Poetry. In K. E. Olsen & L. A. J. R. Houwen (Eds.), Monsters and the Monstrous in Medieval Northwest Europe (pp. 103–122). Leuven: Peeters.
92 Newman, J. L. (2017). Encountering Gorillas: A Chronicle of Discovery, Exploitation, and Survival. Lanham, MD: Roman and Littlefield Publishers.
93 Novaglio, C., Smith, A. D. M., Frusher, S., & Ferretti, F. (2020). Identifying Historical Baseline at the Onset of Exploitation to Improve Understanding of Fishing Impacts. Aquatic Conservation: Marine and Freshwater Ecosystems, 30(3), 475–485. DOI: http://doi.org/10.1002/aqc.3264
94 Nugent, R. (1993). Drums along the Congo: On the Trail of Mokele-Mbemke, the Last Living Dinosaur. Boston, MA: Houghton Mifflin.
95 Olsen, R., & Olsen, K. (2001). Introduction: On the Embodiment of Monstrosity in Northwest Medieval Europe. In K. E. Olsen & L. A. J. R. Houwen (Eds.), Monsters and the Monstrous in Medieval Northwest Europe (pp. 1–22). Leuven, NL: Peeters.
96 Oudemans, A. C. (1892) [2007]. The Great Sea-Serpent. Darke County, OH: Coachwhip Publications (facsimile reprint).
97 Parsons, E. C. M. (2004). Sea Monsters and Mermaids in Scottish Folklore: Can These Tales Give Us Information on the Historic Occurrence of Marine Animals in Scotland? Anthrozoös, 17, 73–80. DOI: http://doi.org/10.2752/089279304786991936
98 Pauly, D. (1995). Anecdotes and the Shifting Baseline Syndrome of Fisheries. Trends in Ecology and Evolution, 10(1), 430. DOI: http://doi.org/10.1016/S0169-5347(00)89171-5
99 Paxton, C. G., Knatterud, M. E., & Hedley, S. L. (2005). Cetaceans, Sex and Sea Serpents: An Analysis of the Egede Accounts of a ‘Most Dreadful Monster’ Seen off the Coast of Greenland in 1734. Archives of Natural History, 32(1), 1–9. DOI: http://doi.org/10.3366/anh.2005.32.1.1
100 Paxton, C. G. M., & Naish, D. (2019). Did Nineteenth Century Marine Vertebrate Fossil Discoveries Influence Sea Serpent Reports? Earth Sciences History, 38(1), 16–27. DOI: http://doi.org/10.17704/1944-6178-38.1.16
101 Pimm, S. L. (2007). Africa: Still the ‘Dark Continent’. Conservation Biology, 21(3), 567–569. DOI: http://doi.org/10.1111/j.1523-1739.2007.00697.x
102 Razafindrakoto, Y., Andrianariveto, N., Cerchio, S., Rasoamananto, I., & Rosenbaum, H. (2008). Preliminary Assessment of Cetacean Incidental Mortality in Artisanal Fisheries in Anakao, Southwestern Region of Madagascar. Western Indian Journal of Marine Science, 7(2), 175–184. DOI: http://doi.org/10.4314/wiojms.v7i2.48275
103 Regal, B. (2012). Richard Owen and the Sea-Serpent. Endeavour, 36(2), 65–68. DOI: http://doi.org/10.1016/j.endeavour.2011.12.001
104 Relaño, F. (2004). Paradise in Africa: The History of a Geographical Myth from its Origins in Medieval Thought to its Gradual Demise in Early Modern Europe. Terrae Incognitae, 36(1), 1–11. DOI: http://doi.org/10.1179/008228804790802200
105 Rick, T. C., & Erlandson, J. M. (Eds.). (2008). Human Impacts on Ancient Marine Ecosystems: A Global Perspective. Berkeley: University of California Press. DOI: http://doi.org/10.1525/9780520934290
106 Riley, M. J., Harman, A., & Rees, R. G. (2009). Evidence of Continued Hunting of Whale Sharks Rhincodon typus in the Maldives. Environmental Biology of Fishes, 86(3), 371–374. DOI: http://doi.org/10.1007/s10641-009-9541-0
107 Ritvo, H. (1998). The Platypus and the Mermaid and Other Figments of the Classifying Imagination. Cambridge, MA: Harvard University Press.
108 Ritvo, H. (2010). Noble Cows and Hybrid Zebras: Essays on Animals and History. Charlotte, VA: University of Virginia Press.
109 Ruddiman, W. F., Ellis, E. C., Kaplan, J. O., & Fuller, D. Q. (2015). Defining the Epoch We Live In. Science, 348(6230), 38–39. DOI: http://doi.org/10.1126/science.aaa7297
110 Saenz-Arroyo, A., Roberts, C. M., Torre, J., Carino-Olvera, M., & Hawkins, J. P. (2006). The Value of Evidence about Past Abundance: Marine Fauna of the Gulf of California Through the Eyes of 16th to 19th Century Travellers. Fish and Fisheries, 7(2), 128–146. DOI: http://doi.org/10.1111/j.1467-2979.2006.00214.x
111 Saunders, R. A. (2018). Reimagining the Colonial Wilderness: ‘Africa’, Imperialism and the Geographical Legerdemain of the Vorrh. Cultural Geographies, 26(2), 177–194. DOI: http://doi.org/10.1177/1474474018811669
112 Scott, A. (2020). The ‘Missing Link’ Between Science and Show Business: Exhibiting Gorillas and Chimpanzees in Victorian Liverpool. Journal of Victorian Culture, 25(1), 1–20. DOI: http://doi.org/10.1093/jvcult/vcz015
113 Six, A. L., & Thompson, H. (2016). From Hideous to Hedonist: The Changing Face of the Nineteenth-Century Monster. In A. S. Mittman & P. J. Dendle (Eds.), The Ashgate Research Companion to Monsters and the Monstrous (pp. 237–256). New York: Routledge.
114 Stratton, J. (2020). Zombie Trouble: Zombie Texts, Bare Life, and Displaced People. In J. A. Weinstock (Ed.), The Monster Theory Reader (pp. 403–421). Minneapolis, MN: University of Minnesota Press. DOI: http://doi.org/10.5749/j.ctvtv937f.24
115 Szabo, P., & Hedl, R. (2011). Advancing the Integration of History and Ecology for Conservation. Conservation Biology, 25(4), 680–687. DOI: http://doi.org/10.1111/j.1523-1739.2011.01710.x
116 Tsing, A., Swanson, H., Gan, E., & Bubandt, N. (Eds.). (2017). Arts of Living on a Damaged Planet: Ghosts and Monsters of the Anthropocene. Minneapolis, MN: University of Minnesota Press.
117 van Druzer, C. (2016). Hic sunt dracones: The Geography and Cartography of Monsters. In A. S. Mittman & P. J. Dendle (Eds.), The Ashgate Research Companion to Monsters and the Monstrous (pp. 387–436). New York: Routledge.
118 van Niekerk, L. K. (2011). Marine Fish Exploitation During the Middle and Stone Age of South Africa. Azania: Archaeological Research in Africa, 46(3), 392–399. DOI: http://doi.org/10.1080/0067270X.2011.609610
119 van Weerden, J. F., et al. (2013). A Meta-Analysis of Steady Undulatory Swimming. Fish and Fisheries, 15(3), 397–409. DOI: http://doi.org/10.1111/faf.12022
120 Vegter, A. C., et al. (2014). Global Research Priorities to Mitigate Plastic Pollution Impacts on Marine Wildlife. Endangered Species Research, 25, 225–247. DOI: http://doi.org/10.3354/esr00623
121 Walsh, M. T., & Goldman, H. V. (2020). Cryptids and credulity: The Zanzibar Leopard and Other Imaginary Beings. In S. Hurn (Ed.), Anthropology and Cryptozoology: Exploring Encounters with Mysterious Creatures (pp. 54–90). New York: Routledge.
122 Weinstock, J. A. (2016). Invisible Monsters: Vision, Horror, and Contemporary Culture. In A. S. Mittman & P. J. Dendle (Eds.), The Ashgate Research Companion to Monsters and the Monstrous (pp. 275–292). New York: Routledge.
123 Weinstock, J. A. (Ed.). (2020). The Monster Theory Reader. Minneapolis, MN: University of Minnesota Press. DOI: http://doi.org/10.5749/j.ctvtv937f
124 Westrum, R. (1979). Knowledge About Sea Serpents. In R. Wallis (Ed.), On the Margins of Science: The Social Construction of Rejected Knowledge (pp. 145–169). Newcastle-under-Lyme: University of Keele.
125 Wong, K. (2012). Rise of Humans 2 Million Years Ago Doomed Large Carnivores. Scientific American, 25 April.
126 Wood, G. D. (2016). Frankenstein, the Baroness, and the Climate Refugees of 1816. Public Domain Review, 15 June. DOI: http://doi.org/10.1086/TWC48010003